The DENNISON Patrilineage 2 DNA Project
DENNISON DNA Patrilineage 2
The Recent Virginia Origins
The ancestors of most, if not all, of the American members of this DENNISON patrilineage appear to have settled in Pittsylvania County, Virginia, by the late 1700s, and migrated from there from 1800-1830 to clusters of westerly Kentucky and Tennessee counties. Even though Pittsylvania lies just to the SE of the Blue Ridge mountains, and just outside the Valley of Virginia where the Scotch-Irish DENNISONs of Patrilineage 1 settled, the yDNA results shows that these two groups of Virginia DENNISONs are completely unrelated.
As it turns out, although the PittsylvaniaCo DENNISONs originally appeared to constitute a single extended family, they actually represent two different patrilineages: besides this DENNISON Patrilineage 2, which all appears to descend from James DENNISON of PittsylvaniaCo, there is also DENNISON Patrilineage 6 which descends from Benjamin and (probably also) Richard DENNISON of PittsylvaniaCo, for which I have yet to construct a project page. The evidence for this is complicated, and I have addressed it thoroughly in the set of reports for which links are provided under “Other Genealogical Resources & Evidence” in the upper left navigation panel of this page. Everything that I had earlier written about Patrilineage 2 in the paragraphs and sections below, however, still stands.
It’s highly unlikely, prima facie, that the PittsylvaniaCo DENNISONs were Scotch-Irish, for a number of reasons, beginning with the fact that most British DENNISONs are English, not Scottish, thus the vast majority of American DENNISONs have English, not Scotch-Irish roots. However, for reasons that I will develop below, I do not think that the ultimate ancestry of this patrilineage was English.
Second, Pittsylvania County was overwhelmingly settled, not by the Scotch-Irish whose initial settlements were beyond the mountains, but by tidewater Virginians slowly trickling west. This was true of Pittsylvania's parent county, Halifax, to the east, and of its child county, Henry, to the west. And the same east to west pattern applied to the next northern tier of counties, from Lunenburg, to Bedford, to Franklin County (where I happen to live). Furthermore, these early settlers of Virginia’s piedmont, and “south side”, continued west mostly in the wake of the Scotch-Irish, progressing west to Tennessee and Kentucky a decade or two after them.
Third, the early settlers of these cismontane counties, including these DENNISONs, do not, by and large, bear Scotch-Irish surnames, nor are their given names confined to the restrictive Scottish (and Scotch-Irish) set, and they don’t appear to observe the customary Scottish and Scotch-Irish onomastic (child-naming) pattern either (see my definitive paper on Scottish naming practices, including the Appendix showing the frequencies of the 20 most popular given names).
At least some of the Pittsylvania DENNISONs came from Maryland
DENNISON Surname Project member, Kathy-52 (of the DENNISON Patrilineage 6 Project) has found the key evidence that links at least her ancestor, Benjamin of BarrenCoKY and PittsylvaniaCoVA, back to Maryland—specifically, a pair of matching records just a couple of days apart for his marriage to Drucilla Pearce—the name “Drucilla”, of Benjamin’s wife, having followed Benjamin, first to PittsylvaniaCo, and then BarrenCoKY, where this couple ended up. The first of these records, dated 26Oct1779 is in PrinceGeorge'sCoMD, and the second, dated 28Oct is in MontgomeryCoMD. It looks as though Benjamin obtained his license in PrinceGeorge’sCo where he lived, but married in MontgomeryCo, probably where he went to church. Thus, he, and presumably other DENNISONs of his extended family, probably lived in PrinceGeorge’sCoMD, somewhere along its extensive border with MontgomeryCo. Also, Benjamin of PittsylvaniaCo’s oldest son was named Zachariah, and there was another marriage record for a Zachariah in PrinceGeorgeCoMD, in 1777, less than two years before Benjamin’s—quite likely this was for an older brother.
There are apparent problems with this evidence, in the form of the widely variant surname spellings in first these MD marriage records, and then in PittsylvaniaCoVA, but after my own now exhaustive research of the underlying primary records, I’ve dealt with this and the other issues that complicate the story of the PittsylvaniaCo DENNISONs, and have presented my conclusions and arguments in the set of linked reports under the head “Other Genealogical Resources and Evidence”, opposite.
Although it may seem a bit of a stretch that these DENNISONs of central Maryland, on the Chesapeake Bay opposite northern Virginia, should have leapt directly to PittsylvaniaCoVA on the border with North Carolina, Benjamin, one of the three PittsylvaniaCo patriarchs, his conjugal family, and scions of the other patriarchs, later made an even greater leap from PittsylvaniaCo to BarrenCo and other south central KY counties. The distance of this latter leap was even greater, while the due west direction was more typical of early American migrations, because in an era when virtually every family engaged in at least part time farming and pasturing, sticking to familiar climates and growing seasons was a prime desideratum.
The deviation from this rule that drove most of the early cismontane Virginians, the vast majority of whom (or their ancestors) were brought to the Chesapeaks and other tidewater counties as indentured servants, was due to the major obstacle of the mountains themselves, and the menace of the unknown wildnerness that lay beyond. When these early Virginia importees were able, subsequent to their term of service, to think of affording their own land (a readiness that was sometimes measured in generations), they were obliged to migrate to whatever counties of VA or NC then constituted the inner cismontane frontier and still offered desirable cheap and unoccupied land. And in the 1770s, PittsylvaniaCo (created in 1767 from HalifaxCo) was one of those counties.
That these DENNISONs should have been in Maryland in the first place nicely dovetails with the discovery, some years ago now, that the ancestry of the Pittsylvania patriarch James was ultimately Scandinavian, and that there’s a strong possibility that his immigrant ancestor was an immigrant to the early (1638) New Sweden colony on the Delaware River, less than 100 miles north of the border between PrinceGeorge's and Montgomery Cos, MD—a subject that I shall take up next. And although Y-Chromosome DNA testing of descendants of the patriarchs James and Benjamin has shown that the former belonged to DENNISON Patrilineage 2, while Benjamin (and probably Richard) descend from DENNISON Patrilineage 6, I nonetheless think it likely that these two were related through the maternal line, as I've explained in a section of my Three Patriarchs paper.
The Ultimate Scandinavian Origins of DENNISON Patrilineage 2
As it turns out, the ultimate roots of DENNISON Patrilineage 2 lie in southern Scandinavia, with several lines converging on the area around Sweden’s principal modern port of Gothenburg. There is also a possibility that the line was originally Norwegian.
Lois-10, the only one of the Amercian Patrilineage 2 Project members who has even tested to 67 markers, has FTDNA-reported matches to two foreigners: Englishman Lee Daniels (member 11), who has traced his ancestry back to 18th century Buckinghamshire, and Simo Roman (member 33), who is a Finn with ancestors who trace back to 18th century Skaraborg County, Sweden, which was recently merged with adjacent Gothenburg. And Lee and Simo in turn have reported matches to Norwegian, Petter Vennemoe (member 35), and to a cluster of men surnamed LONGACRE who have solid descents from one Andrew Peterson Långåker born 1657 in Goteborg, Sweden, who apparently emigrated to the New Sweden Colony on the Delaware.
Petter has traced his Norwegian ancestry even farther back than the LONGACREs, to one Sivert [Olsson?] born say 1590 and a resident of the farm of Skarebøl, Akershus County, a large rural farming community that lies just to the west of Oslo, and to Sivert’s son, Svend, who took up residence in the adjacent farm of Bøen, also in Akershus. Akershus borders Sweden to the east, and lies about 200 miles from the port of Gothenburg overland, though the connection from Oslo to Gothenburg by sea would probably have been more convenient in the 16th and 17th centuries.
Scandinavian Surnames, Bynames, and Patronymics
The LONGACRE surname presumably derives from the Swedish Långåker (“long field” or phonetically “long acre”): at least Reaney & Wilson’s etymological Oxford Dictionary of English Surnames shows no English derivation for the surname. And Petter Vennemoe, the Norwegian member of this patriineage, has found that there was a Swedish farm near Gothenburg named Långåker. Modern Americans might be inclined to say, well, that’s interesting, but so what: certainly it’s possible that the New Sweden family could have hailed from there, but after all, most American and British surnames are derived from personal or occupational names, and only a small minority are locative in origin—derived from place names.
Scandinavian personal names and surnames, however, follow a radically different pattern from what Americans are used to. In the first place, virtually all Americans have had surnames from the time they stepped off the boat—permanent, hereditary, surnames that were passed down by the father to his children. And in England, from whence the largest contingent of American immigrants came, such surnames have been the rule since the 16th century and before. However, in Scandinavia, Russia, and a few other areas, permanent hereditary surnames didn’t become general until as late as 1900.
Instead, individuals with the same given name were differentiated through bynames that applied solely to them, and expired when they did. The most common form, and system, of bynames was the patronymic—a temporary surname derived for a child from its father’s given name (thus Peter, the son of Anders, was known as Peter Andersson). Until 1900 or so, virtually all Scandinavians were known primarily by their given name and patronymic, but as rural populations, and commercial intercourse, grew from the early centuries of scattered settlement, the relatively small stock of Scandinavian given names, each with its derivative patronymic, proved insuffient to create for each person a more or less unique name and identity. This problem was endemic throughout much of Eurasia, and it was solved in various ways, but in Norway, and to a lesser extent in Sweden and Finland, most individuals acquired, at least implicitly, as an additional byname, the name of the farm (or farming district) that they resided within.
Thus, the full names of Petter’s earliest known ancestors were Sivert [Olsson? or Olsen?] of Skarebøl, and his son, Svend Sivertsen of Bøen, or just Svend Sivertsson Bøen. And when a great-great grandson of Svend finally removed to a different farm, he became known as Hans Bergersen, of Kongtorp.
And when Anders Petersson of the New Sweden colony (the son of Peter Andersson originally of Gothenburg, Sweden) chose LONGACRE to be the hereditary surname for his American line, it is highly likely that he would have used his family’s farm name, and that his father and other ancestors and collaterals should have had their roots in the large Swedish farm, Långåker, near Gothenburg, before they were lured to town to participate in the New Sweden adventure.
A Theory of Convergence for this Patrilineage to a Most Recent Common Ancestor
Besides finding a likely area of origin for the ancestors of his LONGACRE cousins, Petter has also proposed a theory to account for the convergence of his line with theirs, and with it, a probable MRCA for the patrilineage as a whole. His theory is predicated on the circumstance that the patronymic, Olsson, that he had independently projected for his probable earliest known ancestor, Sivert of Skarebøl (born say 1590), is the same as the patronymic of the LONGACRE’s earliest known ancestor, Anders Olsson of Gothenburg (born say 1585). And this in turn raises the possibility that these two were brothers and sons of the same father, named Ole.
There are, of course, serious objections to this theory. In the first place, Ole has always been one of the most popular Scandinavian given names, and there are innumerable Oles everywhere. And in the second, the putative sons of these men lived in two different countries, Norway and Sweden.
However, with respect to the latter objection we might observe that the Swedish LONGACRE family was evidently one highly prone to migrate away from their putative local landed area of origin—first to the Gothenburg, and then, almost at the earliest opportunity, to New Sweden, in the New World. Moreover, not long after that passage, perhaps, another descendant of this line who was ancestor to patrilineage member, Lee Daniels, emigrated to England. Given this family history, it is only a small stretch to suppose that the son of a farmer of Långåker, near Gothenburg, should have heard of better opportunities 150-200 miles to the north, just across the border into Norway, and resolved to seek them out. There is no direct evidence that any such thing happened, but it is a perfectly plausible scenario nonetheless.
With respect to the innumerable Oles, we need to avoid anachronism and adjust our thinking to a period when the population of Scandinavia was sparse. Whereas the Swedish population today is around 9.5 million, and the Norwegian about 5 million, interpolating from some early population estimates the population of Sweden about 1590 may have been roughly 700,000, and of Norway, 300,000. Meanwhile, given that the vast majority of modern Scandinavian surnames are derived from patronymics, we can work up a rough estimate of the relative frequency of the given name Ole in Sweden and Norway: about 9% of Norwegians were named Ole, and 5% of Swedes. The product of these estimates yields about 27,000 Oles in all of Norway, and 35,000 in all of Sweden.
We also need to take into account the distribution of this population. Today, a large percentage of Scandinavians live in the big cities and their suburbs (Stockholm and Gothenburg; Olso and Bergen) but things were vastly different in 1600. Although the roots of the city of Oslo/Christiana extend back to the 11th century, the port of Gothenburg wasn’t founded until 1621. The population of Oslo in 1349, just before the Black Death struck, has been estimated at 3,000, but historians have estimatated that that figure had been cut in half by 1400, and, judging by other areas, it would have taken a couple of centuries for it to recover. Meanwhile, Bergen became Norway’s principal port, as part of the Hanseatic league network.
Thus, Scandinavia around 1600 was almost entirely rural, with its population evidently widely scattered, and the denizens of the local farming districts specifically relavant to the Scandinavian ancestors of this patrilineage, are to be measured in the hundreds. Under these circumstances the number of Oles becomes manageable, the number of those with sons named Anders and Sivert even more so.
Finally, although besides these circumstantial considerations, no direct evidence has yet been found linking the New Sweden LONGACREs with Långåker farm, nor any evidence that Petter’s earliest known ancestor, Sivert, was a close relative of the Swedish LONGACRE family, given the extremely tight DNA relationship between Petter and LONGACRE descendants today (Genetic Distance of 1 at 67 markers), it is appropriate that we theorize aggressively to find a possible convergence of their lines in a common ancestor.
In fact, FTDNA’s TIP calculator (adjusted for no known common ancestors for at least 8 generations) says that there's a 38% chance that Petter’s earliest known ancestor Sivert [Olsson?] was also the father of the LONGACRE’s earliest known ancestor, Anders Olsson, and a 76% chance that their lines converge within two generations back from Anders, and since patronymic bynames typically change every generation, with the given name, the most likely hypothesis to account for this data is that these two were sons of the same Ole. What is needed is some intensive research into both the specific records and the history of Långåker and the Gothenburg area of Sweden, and for the Norwegian farms Skarebøl and Bøen to the east of Oslo, near the Swedish border, for the period 1525-1675. Out of this it may be possible to construct a scenario to explain how these two putative brothers ended up in these two different, though not particularly remote, places. I would not foreclose the possibility either, that this patrilineage was originally Norwegian, though I suspect that these distinctions of nationality and ethnicity that we moderns are prone to make are probably somewhat anachronistic as well.
Thumbnail Ancestries of DENNISON Patrilineage 2
What is known of these ancestries may be summarized thus:
(1) Simo Roman has traced his surname patriline back to one Sven Larsson ROMAN, born in Skaraborg County (near the port of Gothenburg) about 1811, and rather remarkably, he has been able to push his patronymic ancestry back another three generations before the adoption of an hereditary surname in his line to one Anders Larsson, born say 1695, though it isn’t known where. However, Anders’s son, Olof Andersson, is known to have died in Söne, Skaraborg County, Sweden, not far from Gothenburg, Sweden’s most important port, so that marks one anchor point for this patrilineage.
The proliferation of surnames here is explained by the fact that prior to the 1800s, the surnames of virtually all Swedes were formed from the (single) given name of their father—the suffix “-son” being used for sons, “-dotter” for daughters. This was essentially the same patronymic system used by the Russians, and for that matter, Welsh, and in all of these areas permanent hereditary suranames didn’t become the rule until well into the 1800s.
(2) The two LONGACREs (who aren’t yet members of the project) descend from one Peter Andersson, an immigrant to the American colony of New Sweden, on the Delaware River, who was himself the son of Anders Olsson of Gothenburg, Sweden, born say 1585. This provides the earliest known anchor point for this patrilineage.
The first colonists arrived from Gothenburg late March 1537/8, under the guidance of the Dutch adventurer, Peter Minuit. The LONGACRE ancestor, Peter Andersson, is thought to have emigrated in 1639 (though I have not myself, seen the evidence for this), and to have shuttled back and forth several times before settling finally in Kingsmessing, near the confluence of the Delaware and the Schuylkill Rivers (now a district of SW Philadelphia). Peter’s son, Anders Petersson, was the first to adopt the permanent hereditary surname LONGACRE (Swedish Långåker), which begins to be associated with his name in the records in 1693. The colonists of New Sweden were among the very first Swedes to have adopted permanent hereditary surnames, as part of their assimilation into English culture.
Significantly, both Simo and his LONGACRE cousins have traced their ancestry back to the vicinity of Gothenburg (Swedish Göteborg), Sweden’s major port. Simo’s earliest ancestors resided in the nearby county of Skaraborg (see this map of the Swedish counties). The specific location of the LONGACRE ancestors is unknown, though it places the line in the Gothenburg area as early as the mid-1600s.
{kind=link}
(3) Lee DANIELS, who is English, has traced his DANIELS line back to Thomas DANIEL[S] born say 1762, probably in Buckinghamshire, near London, Britain’s most important port. Considering the mutational pattern of Lee’s DNA, it’s likely that his line branched off the main trunk of the patrilineage tree very early, some time in the early 1600s, and a near ancestor of Thomas, perhaps his father, probably emigrated to London from Gothenborg some time between 1650-1750.
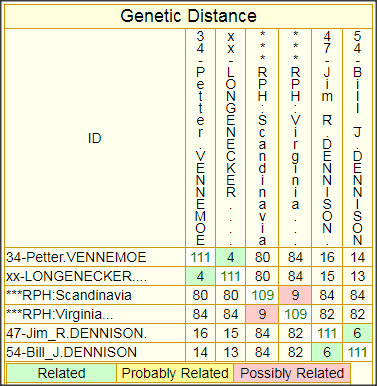
(4) Strictly speaking, member Terrance Nolan (D-45) probably falls just outside this DENNISON Patrilineage 2. Although he comes up as “Possibly Related” to member Simo Roman on the 37-marker GD chart, and improves to “Related” on the 67-marker chart, also picking up a “Probably Related” to Lee Daniels (D-11) of England, his haplotype diverges significantly when expanded to 111 markers. When I compare Terrance’s 111-marker result to that of Norwegian member Petter Vennemoe (D-35), and also to the one LONGENECKER who has extended to 111 markers, Terrance comes short, with a GD=13 relationship (“Unrelated”) to LONGENECKER, and he’s only GD=15 to Petter.
However, I’ve accepted Terrance as a member because his Irish ancestry may shed some light on the deep ancestry of this patrilineage. A GD of 13 projects to a MRCPA with the Scandanavian-rooted ancestors of DENNISON Patrilineage 2 born about the 13th century, give or take a couple of centuries. That puts this MRCPA on the near cusp of the Viking Diaspora, and it seems unlikely that Terrance’s ancestor would have migrated to England during the 14th century—the century of the Black Death—during and after which most Englishmen acquired surnames—English surnames. NOLAN is a distinctively Irish surname that seems to have originated in County Carlow in the SE of Ireland and spread out from there in all directions, but that doesn’t tell us much because so many Irishmen only acquired surnames centuries later. If we can get some other project members to extend to 111 markers, the relationship of Terrance’s line to that of the DENNISON Pat2 line proper, may become clearer.
(5) All the other DENNISON Patrilineage 2 members appear to trace their ancestry back to Pittsylvania County, in south-central Virginia (or possibly to its parent county of Halifax) by the late 1700s, although for only one member, Julie-08, can we now make a specific connection to one of these PittsylvaniaCo families: my recent compilaton and analysis of all the DENNISON entries in the annual tax records of PittsylvaniaCo from 1782-1812, makes it highly probable that Julie’s previous earliest known ancestor, William, who married Jenny Barron in PittsylvaniaCo, was the son of James DENNISON/DONALDSON of PittsylvaniaCo.
Meanwhile, new DENNISON Surname Project member, Kathy Warren Baker (Kathy-52), has traced her ancestry credibly back to Benjamin DENNISON of PittsylvaniaCo, and from there back to Maryland. And since all the indications but one, suggest that James, Benjamin, and a third Pittsylvania DENNISON paterfamilias, Richard, were brothers, James presumably came from Maryland too. I have throughly developed the strong circumstantial evidence in favor of this presumption in my new analytical report on the DENNISONs of PittsylvaniaCo, a link to which, along with other supporting evidence will be found under “Other Genealogical Resources & Evidence” in the top left navigation panel of this page.
However, that one contraindication that James and Benjamin were brothers (or at least full brothers) is fatal: their two credible descendants, Julie-08 and Kathy-52, belong to different DENNISON patrilineages, and they have no common ancestor for at least 2000 years! Clearly an NPE (Non-Paternity Event) has occurred in one of their DENNISON ancestries, but we can pretty well dismiss the possibility that this happened downstream of PittsylvaniaCo, because both have at least two patrilineage cousins who can trace back to different progeny of PittsyvaniaCo principals—though none can as yet actually connect to their senior PittsylvaniaCo ancestor. The resolution to this paradox is that James and Benjamin were probably half brothers, while Richard was presumably a full brother of one of them, most likely Benjamin, considering Richard’s close proximity to Benjamin in the county tax records that I’ve just compiled and analyzed. However, we need to test a patrilineal descendant of Richard to know this for sure.
Finally, given that DENNISON Patrilineage 2 is Scandinavian in its ultimate origin with patrilineage cousins named LONGACRE, and the fact that the area of Maryland where the PittsylvaniaCo DENNISONs came from is only about 100 miles SW of the 1638 New Sweden settlement on the Delaware River, the only important Swedish emigration to America before the 19th century, suggests the possibility that they descended from Peter Andersson, the New Sweden colonist and the immigrant ancestor of the LONGACREs, or a close male relative of his who assumed the surname DENNISON and whose descendants drifted down a ways to Maryland. I’ve considered this specific hypothesis in detail in the next section analyzing the DENNISON Patrilineage 2 DNA.
Implications of the DNA Evidence for DENNISON Patrilineage 2:
Additional yDNA Testing is Needed
First, although I’ve tried below to milk what I can out of the yDNA evidence so far, it would be highly desirable to extend to 111 markers the haplotypes of the rest of the current members of DENNISON Patrilineage 2, particularly the American members, in order to look for possible Closer Cousin Cluster (CCC) relationships among them that could be useful in focusing and guiding further research to connect them back to PittsylvaniaCoVA. Extending would also provide a better read on the closeness of relationship between the American DENNISONs and the LONGACREs and other likely descendants of the New Sweden settlement.
As a separate matter, it would also be desirable to try to track down descendants of Richard DENNISON of PittsylvaniaCo and test them to see whether they too belong to Patrilineage 2, or to Kathy-52’s DENNISON Patrilineage 6. Knowing that would help in looking for the NPE (Non-Paternity Event) that must have occurred, probably in Maryland, in the siring of one of the three PittsylvaniaCo DENNISON brothers: James, Benjamin, and Richard.
The Genetic Distance Evidence
To provide an overview of the closeness of these patrilineage interrelationships I have updated the DENNISON Patrilineage 2 67-marker GD matrix for the five 67-marker haplotypes that have surfaced for this patrilineage, as well as the less accurate 37-marker matrix, into which I’ve incorporated all the additional 37-marker LONGACRE haplotypes. The 67-marker matrix shows these DENNISON, DANIELS, LONGACRE, and ROMAN haplotypes are unequivocally “Related ” (in FTDNA’s parlance), and the 37-marker matrix shows that they have many tested LONGACRE cousins, a couple of whom might profit from extending to 67 markers. As a matter of fact, it would be desirable at some point for all the members of this project to extend their haplotypes further to 111 markers, both to look for Closer
In light of these expanded GD matrices, I have adopted Simo ROMAN’s haplotype as the new RPH for DENNISON Patrilineage 2. Simo’s largest GD at 67 markers with any of the others (which is 6), projects to a MRCA (Most Recent Common Ancestor) for all 5 of the patrilineage members who have tested to 67 markers to a man who was born say 1500, while the maximum GD between any of these 67-marker haplotypes is 7, which projects back to a common ancestor born say 1425. However, there is so much slop in these estimates that one needs to make a mental note to append to any such estimates: “give or take a century or two”. Extending all these haplotypes to 111 markers should tighten these loose estimates significantly, bring the variance down to less than a century either way. Now how do these TMRCA estimates, predicated entirely on the mutational probabilities, square with the known genealogy?
The GDs of 6 and 7 separating Englishman Lee DANIELs from his Scandinavian cousins are in line with the idea that his earliest known ancestor, Thomas DANIEL (born say 1762) was a descendant of a Swede who emigrated between 1600-1750 from Sweden’s most important port, Gothenburg, to England’s most important port, London.
Likewise, the GD 6 and 7 matches between Lois, a DENNISON, and her ROMAN and LONGACRE cousins from the area of Gothenburg, Sweden, are consistent with the hypothesis that she and her PittsylvaniaCoVA cousins descend from a fairly close patrilineal relative of Peter Andersson, the ancestor of the LONGACREs, who emigrated from the Swedish port of Gothenburg to New Sweden on the Delaware about 1639. However, it is just as likely, considering her GD of 7 from Englishman Lee DANIELS, that her line, and that of the other Virginia DENNISONs, is a bit more closely related to the English DANIELS line and that they came, or were transported, to Virginia as indentured servants, like the 80% or so of Virginia immigrants during the colonial period—more on this below.
What seems much harder to swallow is the very low GD between Simo ROMAN, a Finn with exclusively Finnish and Swedish ancestry, and his two American LONGACRE cousins, whose direct ties with their country of origin presumably ended with the death of the immigrant Peter in 1679 about 8 generations back, and whose only known son first assumed the surname LONGACRE. Meanwhile, Simo has traced his patrilineal ancestor 7 generations back to a man named Anders Larsson, born say 1695, so supposing, say, that Simo’s earliest known ancestor was a great-grandson of Peter’s father, the expected (or most likely) number of mutations would be 3.6, and the probability that there had been only one mutation would be only 10%.
In fact, from the little I have learned of the LONGACRE ancestry, the perfect match between the two LONGACRE descendants at 67 markers looks suspicious too. These two are supposed to be descended from two different sons of the immigrant Peter’s son, Anders Petersson LONGACRE (1657 - abt Nov1718), and if we guess that it is 9 generations back to Anders, the expected number of mutations is 4 between these two descendants, not 0, for which the probability is less than 2%.
This may all be just a case of long odds results turning up now and then, but the conjunction of two long odds results having a composite probability of just 1 in 500 (.1 x .02 = .002) leads me to consider alternative hypotheses for the respective genealogies here. There seems little room to question whether Simo’s patrilineal ancestry is purely Scandinavian, and there was precious little back migration to Europe for most American immigrants, so that leads one to wonder whether the father of his first ROMAN ancestor may not have been also a father or close collateral of Swedish emigrants to America in the early 1800s, who for whatever reason adopted the LONGACRE surname. These hypothetical later immigrants may, in fact, have been distant cousins of the first LONGACRE of New Sweden, who maintained their extended family ties over the nearly two centuries for which they were separated. The probability of this is increased if it is true that the original New Sweden immigrant, Peter, made several trips back and forth across the Atlantic before finally settling in America for good.
That would be one way of accounting for this anomaly, but perhaps a better way is to back up and look at all seven of the LONGACRE haplotypes at the 37-marker level. We find in the first place a mutation in common for all of them, CDYa=36, that differentiates them from the haplotype of Simo and all the others. Second, the set of seven LONGACRE haplotypes as a whole looks like a typical set of haplotypes descended from a mid to late 17th century American immigrant. The two 67-marker haplotypes that I’ve been presuming all along belong to the two LONGACREs who descend from different grandsons of the immigrant Peter are still anomalous in being identical, but it’s always possible that my presumption is wrong, and even if not, much stranger things than this have happened.
The Fine-Grained Mutational Pattern Evidence
A fine-grained comparison of the specific marker values across the patrilineage indicates that DANIELS and DENNISON share at least one marker value that none of the LONGACREs have: they are DYS447=24, while the LONGACREs are DYS447=25. There are two possible interpretations of this: either DYS447=24 was the original value, and the mutation to 25 occurred after the hiving off of DENNISON and DANIELS, but before the emigration of the LONGACRE ancestor, and was thus inherited by all the tested LONGACREs, or the original value of DYS447 was 25, in which case the mutation to 24, which was inherited by both DENNISON and DANIELs, occurred during the 1-3 generations that their lines ran together before their final separation, probably due to the emigration of one or the other out of Sweden.
Without having a detailed grasp of the LONGACRE genealogy, and being able to evaluate the evidence behind it, it is difficult to choose between these alternatives (or even to be quite sure about the claimed LONGACRE descent from one Andrew Peterson Långåker born about 1657, which the DNA suggests), however, I do note from statistics on DYS447 at the Sorenson Foundation that DYS447=25 is the prevalent value across all haplotypes (3x as common as 24), and that it is also the prevalent value in the LONGENECKER and other R1b projects that I’ve looked at, so I will guess that it is the DYS447=24 value that is the mutation here. In which case it is more likely than not that the DENNISON and the DANIELs line ran together for several generation after diverging together from the main trunk.
Not too many generations, though, because the DENNISON line saw a mutation of the RPH (Simo ROMAN & LONGACRE) marker value of DYS449=31, first to 32 (for the Lois-10 haplotype), then another mutation in the same direction to DYS449=33 (for the other three DENNISON haplotypes), while in the meantime, the DANIELS haplotype at some point mutated in the other direction (losing instead of gaining a repeat) to DYS449=30.
Since it appears that Englishman Lee Daniels’s ancestor emigrated from Sweden some time during the period of Swedish expansion (1600-1750), presumably sans surname, one would expect that the DENNISON line was most likely brought to England by the same ancestor or by a close collateral, who happened to adopt the different surname DENNISON—a surname that I note fits with the preferred Scandinavian patronymic pattern. Of course other scenarios are possible, but they all have to accomodate the fact that the fine-grained DNA evidence suggests that the line that was to become English DANIELS and that which was to become DENNISON ran together for just a few generations during this approximate period. And as I’ve noted above, extending the marker panels of these descendants to 111 markers would probably tighten up the very broad estimates that we are stuck with here.
Finally, I note that the assumption of a Swedish-English origin for the DENNISON line fits best with their emergence in Southside Virginia about the mid-1700s, just when the descendants of original tidewater servants were settling on this inner Virginia frontier, and the given names found in this Virginia family are also strongly suggestive of specifically English origins.
How the History of the DENNISON Branch of the Patrilineage
Might Intersect with Swedish History
If the DENNISONs of Pittsylvania County, in southern Virginia, were originally from Scandinavia, we need an explanation for how they ended up in the backcountry of colonial Virginia, and I believe that there is one ready to hand. First, though, we must consider a bit of Swedish history.
By and large, the emigree Swedes preferred the cold climates they were used to, so they migrated principally to Canada or the northern tier of the US (particularly Minnesota and the Dakotas), mostly in the mid to late 1800s. However, Sweden had expanded its sphere of influence about the same period that the Dutch were feeling frisky and establishing trading colonies in the New World, and the Swedish monarchs sponsored several of their own resettlement projects. But the only one that contributed significantly to American colonial history, was the colony of New Sweden (1638-1655), headed by Peter Minuit (also a director of the Dutch Colony of New Amsterdam), that was planted on the Delaware River before the North American colonial picture had quite gelled (here is another take on New Sweden).
This expedition was organized and launched from the Gothenburg area, and one presumes would have initially been confined to that population, but later waves of settlement were comprised largely of the Forest Finns—mostly ethnic Finns, but also poor Swedes whom the Swedish Crown had originally encouraged to settle in the northern waste areas of Finland, where they practiced slash and burn agriculture, and proliferated beyond the ability of this poor land to support them. It is claimed by LONGACRE descendants, however, that their ancestor, Peter Andersson, came over as early as 1639, so it is unlikely that he was part of the Forest Finn diaspora.
Inevitably, like the Dutch colony of New Amsterdam up the Atlantic coast from New Sweden, this Swedish project was elbowed aside by the much larger tide of English settlers, although the original Swedish settlers, who numbered about 600, were offered compensation by William Penn in the form of a 10,000 acre tract in Pennsylvania up the Schuylkill River called the Manatawny Creek settlement, and it appears that most of the colonists relocated there. Others, however, fanned out south into the Chesapeake tidewater region and merged with the population of tidewater Virginians who gradually pushed the line of settlement out into the Virginia and Carolina piedmont area on the doorsteps of the Blue Ridge Mountains.
It would seem that this history of Swedish colonization dovetails almost perfectly with the likely history of the DENNISON line in America. In particular this article on the breakup of New Sweden provides examples of Swedes filtering down to the Southside Virginia county of Bedford (created 1754 from LunenburgCo) which had a long border with Pittsylvania (where the Virginia DENNISONs first emerge) when that county was created in 1767 from Halifax County. Of course, it is also possible that these Scandinavian DENNISONs of Southside Virginia diffused initially west into the Pennsylvania hinterland, and came to Virginia later, taking the North-South roads paralleling the Blue Ridge Mountains down to BedfordCo and parts south.
Research Implications of the Theories Presented Here
Unfortunately, I am unable to comment on the prospects for further research in Scandinavia because that lies well outside my area of expertise. Nor have I heretofore had occasion to research the colony of New Sweden on the Delaware. However, it is a point in favor of the work that both Simo ROMAN has done, and that the LONGACREs have relied on that their respective patrilines appear to converge on the Gothenburg area in southwestern Sweden, while the DNA tells us that they must converge somewhere.
However, the colonial records of New Sweden, and of Pennsylvania and Virginia, are accessible to us American researchers, in the usual libraries, archives, online resources, and most of all, LDS films. From my correspondence so far with one LONGACRE descendant, I gather that most or all of what is known about the first two generations of their line comes from a pair of books by Peter Stebbins Craig: The 1693 census of the Swedes on the Delaware (1993), and 1671 census of the Delaware (1999). And he has been good enough to send me photocopies of the relevant pages on his ancestors, from which I have culled the material above. Craig cites his sources, both primary and secondary, though most of these resources are at present out of reach due to the COVID-19 closures. One of these sources, though, is the New Sweden Journal, and among the historical journals cited that I recognize is the Pennsylvania Magazine of History and Biography 2:226, so these secondary sources would be a couple of starting points for locating the ultimate, primary sources regarding the American immigrant generation of DENNISON Patrilineage 2.
Lee Daniels, meanwhile, needs to look for records and other material on immigration from Sweden during the period 1650-1750.
DENNISON Patrilineage 2 Descendancies
The patrilineal descendancies posted below closely follow what the members of this project have submitted, and are both plausible and generally backed by evidence. They are also generally consistent with the best published secondary sources . Although there are areas of weakness in these lineages, due to less than exhaustive research, or to inherently sketchy records, a significant and ongoing effort is being made to ensure that what is posted here in abbreviated form represents the best current thinking on these patrilineal descent trees.
Each descendancy begins with the earliest known patriarch of a particular line, and follows the patriline down as far as it is known, and at least to its culmination in a DNA-tested descendant. Since this DNA patrilineage project is focused on tested or testable males of the patriline, these descendancy trees have generally been pruned of daughters or stepchildren of previously married wives. On the other hand, known brothers of project member’s ancestors have been included, and in some cases their patrilineal descendancies have been followed as well, to facilitate the identification of testable patrilineal descendants (living males who in most cases bear the focal surname). Where one of these collateral male lines is known to have petered out (or daughtered out), it will be flagged “no known sons”, NKSons, or words to that effect.
The information provided for each male ALLEN should be just sufficient in most cases to uniquely identify him in the USCensus and other readily available sources. They are not meant to be complete genealogies. The headnotes for each patriarch identify the principal researcher(s) of that line, and often contain links to relevant webpages, compilations, or more complete genealogical material.
For times and places where permanent hereditary surnames were in general use, I have capitalized the surnames only of the wives, to help these stand out. For other populations I have capitalized the permanent hereditary surname the first time it appears in the male line of descent.
Thus, in general, Scandinavian surnames before 1900 were bynames formed from the patronymic root. For example, Lars Andersson, was Lars the son of Anders, whose own byname was a patronymic based lifewise on his father’s given name with the addition of the “-sen” (or Anglicized “-son”) suffix. And besides the patronymic, most pre-1900 Norwegians, and perhaps other Scandinavians too, had as a second byname, the name of the large farms on which they worked, and some of these farms and farm names go back to the 12th century. Thus, the Lars Andersson of my example might be differentiated from others of that name by referring to him more fully as “Lars Andersson, of Bøen”(Bøen is a large farm near Oslo). One might equally well omit the comma, or refer to him as “Lars Andersson Bøen”, but I have interpolated the “of”, and the comma, to distinguish these locative bynames, that might change if their subject changed his/her residence, from the permanent hereditary surnames that came in later, and I have additionally capitalized these permanent surnames the first time they appear, to mark them out.
The data posted below for each male of the patriline comprise (insofar as is known): his date and place of birth, date and place of death, the name(s) of his wife (or wives), and the date and place his marriage(s). The data are presented in a standardized abbreviated format designed to facilitate scanning and CTL-F searching.
Specific dates (where known) are abbreviated to year dates. Dates that aren’t known specifically, or which aren’t backed by uncontroverted evidence are qualified as either approximate (“abt”, “bef”, “aft”, or “by”), or where they are merely guesstimated, by (“say”). Approximated dates imply supporting evidence which merely fails of complete accuracy, while “say” dates are guesstimates based on typical patterns of the time, place, and social group.
Places are abbreviated to the most important jurisdictional place where records are to be found: for most US states/colonies these are counties; for New England, towns. Places should be specified only where there is either direct evidence, or overwhelming circumstantial evidence, that a vital event occurred in that place and none other, and they shouldn’t be over-specified with respect to the actual supporting evidence.
The yDNA-tested male descendants in whom these descendancies culminate are flagged below with the ShortName of the Principal Researcher of their line (the person who represents the descendant’s haplotype), and by the Project# for the membership, for example, Julie-08.
Notes about the ySTR DNA mutations that mark particular lineages have been interwoven below in red text. Notes about upstream mutations distinctive of particular family sub-branches have been inserted in the descendancy at the point at which they begin to be relevant, and where those same mutations have been inherited by tested descendants, this has been noted right after the name(s) of the tested descendant and Principal Researcher of the line. Please note, however, the careful qualifications in these DNA notes. Most inferences drawn from DNA evidence are probabilistic in nature and one needs to keep an open mind about alternative interpretations, just as one does with the genealogy itself.
Doubtful ancestral links or children are outlined in yellow, e.g. |.
invisible writing
invisible writing
-----The conventions that govern the following Patrilineage 2 Descendancies are defined above-----
invisible writing
1-James Dennison/Donaldson of Pittsylvania County, Virginia, born in MD?
Sources: Julie Dennison, Bill_J Dennison, Bob_L Dennison, and John Robb.
1-James Dennison/Donaldson (say 1752 perhaps in MD - aft 1812 [PittsylvaniaCoVA?])
The mutations DYS392=12 & DYS464=15-15-17-17 mark all the known descendants of 1-James.
|--2-William Dennison (abt 1782 PittsylvaniaCoVA - aft 12Oct1860)
| ---m(bond). 4Oct1803 Jenny BARRON (d/o Josiah [& Elizabeth?]), in PittsylvaniaCoVA
The mutations DYS390=23 and CDYb=39 mark all the descendants of 3-Trammel,
and maybe those of father 2-William as well.
| |--3-Trammel Dennison (abt 1807 VA - 1891 GraysonCoKY)
| | ---m. say 1825 Frances CARTER
| | |--4-Henry Clay Dennison (1832 GraysonCoKY - 1890 GraysonCoKY)
| | | ---m. Elizabeth A. WEEDMAN
| | | |--5-Charles William Dennison (1857 KY - 1939 SangamonCoIL) m2. Sarah T. KIPER
| | | | |--6-Clarence E. Dennison (1895 GraysonCoKY - 1965 McLeanCoIL)
| | | | | ---m. Elizabeth COTTON
| | | | | |--7-William Lee Dennison (1944 McLeanCoIL - 2001 IL)
| | | | | | |--8-son of William *** Julie-08 ***
| | |--4-Claiborne Dennison (1835 GraysonCoKY - 1917 GraysonCoKY)
| | | ---m1. 27Sep1854 Sarah L.M. WILSON, in GraysonCoKY
| | | |--5-James Will[iam?] Dennison (27Oct1855 GraysonCoKY - 1900 GraysonCoKY)
| | | | ---m. 3Jan1878 Katherine A. DENNISON, in Grayson CoKy
| | | | |--6-Joseph(“Joe”) Claibourn Dennison (abt 13Sep1885 GraysonCoKY - 1957 GraysonCoKY)
| | | | | |--7-William C Dennison
| | | | | | |--8-William J Dennison *** Bill_J-54 ***
| | | ---m2. 1866 Rose J. STONE, in GraysonCoKY
| | | |--5-Jabez Percival Dennison (1886 KY - 1944 MasonCoIL)
| | | | ---m. 1907 Lula CARTER, in Grayson CoKy
| | | | |--6-Murrell Dennison
| | | | | |--7-Robert Leland Dennison *** Bob_L-26 ***
| | |--4-James Dennison (abt 1838 GraysonCoKY -)
|--2-William Dennison (abt 1782 PittsylvaniaCoVA - aft 12Oct1860)
| ---m(bond). 4Oct1803 Jenny BARRON (d/o Josiah [& Elizabeth?]), in PittsylvaniaCoVA
The mutations DYS390=23 and CDYb=39 mark all the descendants of 3-Trammel,
and maybe those of father 2-William as well.
| |--3-Trammel Dennison (abt 1807 VA - 1891 GraysonCoKY)
invisible writing
|--2-Stephen Denison (1789 VA - 1866 [HendersonCo]TN)
| ---m(bond). 15Oct1813 Elizabeth INGRAM, in PittsylvaniaCoVA
The mutations DYS552=23 marks the one known descendant of 2-Stephen.
| | The putative sons of 2-Stephen appear either in his DecaturCoTN household in the 1850 USCensus,
| | or, with one exception, heading their own households in adjacent HendersonCo.
| | Their number also matches Stephen’s 1830 household in PerryCo, the parent of Decatur.
| |--3-Robert R. Denison (abt Sep1814 VA (VA,VA) - 1908 HendersonCoTN)
| |--3-Byrd Denison (abt 1819 TN - ) - migrated to TX by 1860 where he a son named Curry - father b.VA in 1880
| |--3-M. Denison (abt 1822 -) - probably the carpenter in the 1850 MontgomeryCoTN HH of M. Wilburn
| |--3-Sanford Denison (abt 1825 NC/TN - aft 1Jun1880 [TX?])
| |--3-William H. Denison (abt 1829 TN - aft 1Jun1880 [TX?])
| |--3-A.J. Denison (abt 1830 TN - bef 1870?)
| |--3-Benjamin L. Denison (abt 1831 - aft 1Jun1880 [TX?])
| |--3-David [Gray?] Denison ([1Sep?]1837 - [29Apr1866?])
| | ---m. [Nancy WALLACE?]
| | |--4-John Steven Dennison ([29?]May1860 - [30Mar?]1940)
| | | ---m. abt 1883 Frances [Josephine DUKE?]
| | | |--5-Nicholas David Dennison (21Mar1898 - 10Jul1979 GibsonCoTN)
| | | | ---m. Fredda Lee JENNINGS
| | | | |--6-James Royce Dennison Sr.
| | | | | |--7-James Royce Dennison Jr *** Jim_R-47 ***
| | |--4-Curry S. Dennison (abt Mar1864 - aft 1900)
| |--3-Curry P. Denison (abt Nov1839 TN - aft 19Apr1910 [DecaturCoTN?])
invisible writing
1-Joel Dennison of Virginia, born abt 1808
Sources: Lois Franceschi
Joel’s descendants share the mutations DYS392=12 & DYS464=15-15-17-17
with the known descendants of 1-James of PittsylvaniaCoVA.
1--Joel Dennison (abt 1808 VA -)
|--2-[m. 20Apr1838 Anna JACKSON, in BarrenCoKY]
|--2-James W. Dennison (abt 1839 KY -)
|--2-John Burch Dennison (1840 KY - 1917 ReynoldsCoMO)
|--2-Daniel Albert Dennison (1844 KY - 1912)
|--2-Jeremiah M. Dennison (1849 MO - 1920 IronCoMO)
|--2-Benjamin Franklin Dennison (1852 ReynoldsCoMO - 1934 ReynoldsCoMO)
| ---m. Rebecca Jane STEVENS
| |--3-Elmer Joel Dennison (abt 1905 MO -) m. Virgie Marie HENSON
| | |--4-James Dale Dennison m. Irma Sue ROOD *** Lois-10 ***
(has mutation DYS449-)
invisible writing
1-Anders Larsson of Sweden, born say 1695
Sources: Simo Roman
The surnames of Simo’s ancestors before ROMAN and BERG were patronymics
in the manner of early Russian surnames.
1-Anders Larsson (say 1695 -) m. Elin Andersdotter
|--2-Olof Andersson (say 1725 1798 Söne, SkaraborgCo SWE)
| ---m. 1757 Elin Andersdotter, in Söne
| |--3-Lars Olofsson (1761 Söne, SkaraborgCo SWE 1820 Österlösa, SkaraborgCo, SWE)
| | ---m. Kjerstin Larsdotter
| | |--4-Sven Larsson ROMAN (1811 Örslösa, SkaraborgCo SWE 1891 Skara, SkaraborgCo SWE)
| | | ---m. 1837 Carolina BERG, in Skara, SkaraborgCo
| | | |--5-Ernst Josef Roman (1852 Skara, SkaraborgCo SWE 1911 Österåker, Stockholm SWE)
| | | | ---m. 1884 Matilda Charlotta ODHNER, in Forshem, SkaraborgCo SWE
| | | | |--6-Sven Alfred Roman (1885 Fullösa, SkaraborgCo SWE 1966 Strängnäs, SödermanlandCo SWE)
| | | | | ---m. 1920 Svea Elisabeth SPIIK
| | | | | |--7-Finn Olof Gunnar Roman (1924 Helsinki, FIN 1976 Helsinki FIN)
| | | | | | |--8-Simo Antero Roman *** Simo-33 ***
invisible writing
1-Sivert [Olsson?] of son Skarebøl, Akershus, Norway, born say 1590
Sources: Petter Vennemoe
1-Sivert [Olsson?], of Skarebøl (say 1590 [SWE?] - abt 1660 Høland, Akershus, NOR)
|--2-Ole Sivertsen, of Skarebøl (abt 1615 Høland, Akershus - aft 1689)
|--2-Svend Sivertsen, of Bøen (abt 1617 Høland, Akershus, NOR - abt 1702 Høland, Akershus, NOR)
| ---m. abt 1640 Aloug Pedersdatter, of Bøen
| |--3-Berger Svendsen, of Bøen (abt 1651 - 1723 Høland, Akershus)
| | ---m. abt 1682 Guri Knutsdatter, of Bøen
| | |--4-Svend Bergersen, of Bøen (abt 1683 - 1730 Høland, Akershus)
| | | ---m. abt 1715 Ellen Svendsdatter, of Rakkestad
| | | |--5-Berger Svendsen, of Bøen (1729 - 1800 Høland, Akershus)
| | | | ---m. 1749 Ragnild Jørgensdatter, of Burhol
| | | | |--6-Hans Bergersen, of Kongtorp (1767 - 1847 Høland, Akershus)
| | | | | ---m. 1798 Gunild Marie Svendsdatter, of Skarrebøl
| | | | | |--7-Kristian Hansen VENNEMOE (1812 - 1901 Høland, Akershus)
| | | | | | ---m. 1838 Kari Tostensdatter, of Torp
| | | | | | |--8-Per Vennemoe (1918 Oslo, NOR - 1987 Oslo, NOR)
| | | | | | | ---m. 1948 Chicago, USA Elise Bendixen FALCK
| | | | | | | |--9-Petter Falck Vennemoe *** Petter-35 ***
|--2-Jon Sivertson (abt 1626 -)
invisible writing
1-Thomas Daniel of Buckinghamshire, England, born say 1762
Sources: Lee Daniels
1--Thomas Daniel (say 1762 Bucks ENG - 1828 Bucks ENG) m. Judith STEARS
|--2-Joseph Daniels (1805 Bucks ENG -) m. Sarah TIMS
| |--3-William Daniels (1830 Bucks ENG -) m. Hannah BRAGGINS
| | |--4-Andrew William Daniels (1865 Bucks ENG -) m. Lavinia JONES
| | | |--5-Ernest John Daniels (1897 London ENG - 1951) m. Dorothy Caroline WATTS
| | | | |--6-John Frederick Daniels (1943 London ENG -) m. Joan Ann WINSLADE
| | | | | |--7-Lee John Daniels (1966 London ENG -)
*** Lee-11 ***
(has mutations DYS392+, DYS449---, DYS464, DYS576+, and DYS481+)
|--2-Edward Daniels (1807 -)
|--2-George Daniels (1808 - 1887)
-
-
This project is in no way affiliated with the DNA testing company, Family Tree DNA, or any of its surname projects, even though many of the members of this independent patrilineage project may also be members of the FTDNA DENNISON Surname Project. However, this patrilineage project is affiliated with the independent DENNISON Surname Patrilineages Association (DenSPA), whose umbrella page includes material and evidence pertaining to the surname as a whole, and to its bearers of various DENNISON patrilinages.
Navigating from here
The menu buttons at top right take you to other pages on this site, while the nav panel above targets other points on this page, or brings up other resources (papers I’ve written, and the like). If you find yourself lost, the browser BACK button will take you back to where you were (some people also have a convenient BACK button on their mouse, right under their thumb). Or hitting the HOME key of your keyboard will take you back to the top of this page where you are now.
Some Key Terms: CCC, haplotype, haplogroup, NPE, patrilineage, RPH.
DENNISON Patrilineage 2 Directory of Researchers
Active researchers of this DENNISON patrilineage are shown below. Those with highlighted names may be e-mailed by clicking on their names, and their posted descendancies may be viewed by clicking on their highlighted Patrilineage Project#s. Satellite members of the project are listed immediately after the principal researcher for the sublineage they are interested in, and their names are preceded by a dash. Where the person tested is not also the principal researcher, the former’s name appears under the latter’s, in parentheses.
| Proj# | “Handle” | Researchers
(Test Subject) |
yDNA Test Panels |
| D-54 | Bill_J | William J. Dennison | 111 |
| D-09 | Dr. Bob | Dr. Bob Dennison | 37 |
| D-26 | Bob_L | Robert Leland Dennison | 37 |
| D-47 | Jim_R | James Royce Dennison, Jr | 111,Y700 |
| D-08 | Julie | Julie Dennison | 37 |
| D-11 | Lee | Lee John Daniels | 67 |
| D-10 | Lois | Lois Franceschi (Dale Dennison) |
67 |
| D-35 | Petter | Petter Falck Vennemoe | 111 |
| D-33 | Simo | Simo Antero Roman | 67 |
The Deep Ancestry of DENNISON Patrilineage 2:
Haplogroup: R1b -> R-M269 -> L21 -> Z253 -> BY4065
A man’s yDNA may be classified according to a deeper, broader, ancestral schema by testing ySNPs instead of ySTR markers. These ySNPs are unique point mutations to the yChromosome that occur so much less frequently than ySTR mutations that they are of no use in sorting people into patrilineages, but by the same token they are ideal for sorting patrilineages into the various haplogroups and subclades of the broad human population. And by correlating the haplogroup subclades of both modern descendants and of their ancient ancestors (by testing their remains) with geographic population flows, and with archaeological evidence signifying cultural groupings, the whole broad history of homo sapiens is gradually being reconstructed.
DENNISON Patrilineage 2 falls into the broad European haplogroup R-M269, and its subordinated haplogroup R-L21 on the YCC-based haplotree built by FTDNA. FTDNA has so far recognized and made available for testing hundreds of SNPs subordinate to L21, in a branching tree more than 20 levels deep in some lines, and project member Petter Vennemoe has paid for the extra ySNP testing to take DENNISON Patrilineage down to just about the lowest (most recent) level recognized in the FTDNA Haplotree, R-BY4065. The ISOGG organization also maintains an R-tree, but it’s branching ends with the ySNP Z253.
Here’s the complete chain of ySNP branches in the FTDNA Haplotree linking R-M269 down to its subordinate subclased R-L21
M269 -> L23 -> L151 -> P312 -> Z290 -> L21
and here’s the continuation of the tree from L21 down to the lowest (most recent) current level, BY4065
L21 -> DF13 -> ZZ10_1 -> Z253 -> BY4086 -> FGC17432 -> A494 (a.k.a. BY60)
-> S7898 -> Z18132 -> BY157 -> BY4065:
Specialized projects have been organized at some of the nodal levels of this branching tree, and those who have paid for the extra testing, and whose results classify them as at or subordinate to those nodal levels are encouraged to join these projects to help work out the corresponding human population history. The idea is that by bringing this branching ySNP tree down into the range of genealogical time, and noting the geographic areas where the earliest documentable ancestors came from, human population history can be mapped onto geography, which would certainly benefit genealogists who know, from ySTR DNA testing only that they belong to a particular genealogical patrilineage.
In theory if even one member of a ySTR-based genealogical patrilineage project has been tested down to a recent node on the ySNP haplotree, and one or two members can trace their ancestry far enough back to localize it to the same geographic area, all the tested members of that ySTR patrilineage will know where to look for their deepest ancestral roots.
To my knowledge, the most recent nodal branch point for the DENNISON Patrilineage 2 descendancy for which a ySNP project has been organized is R-Z253, and any interest members of our project may wish to apply to join this R-Z253 project. The goal of most of these projects is to provide an umbrella for the working out of subclades and the assignment of their members to these subclades, while publishing both their haplotypes and the surnames these haplotypes correspond to. Sometimes, just by studying the surnames collected into these subclades, amplified in some cases with geographic information, one can get an idea of where the subclade originated.
Since all members of a ySTR patrilineage project share essentially the same Y-Chromosome, with but minor variations, there is no reason for more than one member of a ySTR patrilineage project to pay for additional ySNP testing, but it can be beneficial for all if at least one such member does.
The L21 Subclade
The L21+ subclade of R1b is very close to the Western Atlantic Modal Haplotype (WAMH) that is found all along the European seaboard from Portugal to Scandinavia, and R1b is by far the most common subclade in the British Isles. The L21 SNP mutation is thought to have originated in southern or Alpine Germany about 4000 years ago, and spread north and west from there into the European continent, and thence to Britain, and especially Ireland and Scotland, where, like the Celtic culture, it predominates. However, it's a mistake to identify L21, per se, with the Celtic, or Hallstatt culture that it long predates. Here is a map showing the spreading out and differentiation of R1b over the last 20,000 years. In a new book (published May2011), “The Scots: A Genetic Journey”, by Alistair Moffat and Jim Wilson, the authors have identified the SNP S145, which has always been found so far associated with L21, as a marker for the ancient Pictish culture of Scotland, but as the R1b distribution map indicates, L21 is widely distributed throughout Britain, and takes it’s origins many thousands of years ago on the NW European continent.
At the rate progress is being made in the reconstruction of human history through DNA analysis, before too many more years have passed, I expect that some of these subclades will be brought down into genealogical time, and may even become a shortcut means of identifying patrilineages. In the meantime, you can read more about haplogroups and their distribution across the continents at this site, and there are already efforts to further break down the broad L-21* group by means of haplotype analysis on this DNA Forum thread (you need to register with the Forum, though, to access it).
DENNISON Patrilineage 2 Project NEWS
18Nov2020
The newest member of the DENNISON DNA Surname Project, Kathy Warren Baker (Kathy-52), has injected new life, not only into the DENNISON Patrilineage 6 Project into which her 111-marker yDNA test results classify her, but also into this DENNISON Patrilineage 2 project. As it turns out, Kathy, like Julie-08 of this Patrilineage 2 Project, credibly traces her DENNISON ancestry back to Pittsylvania County, Virginia.
This is rather startling, because, until now, it has been presumed that all the PittsylvaniaCo DENNISONs (who begin to appear about the same time in this inner frontier of Virginia county) were of the same extended family, and the same patrilineage.
This new information has in turn, provoked me to finally mine and process the one major untapped body of PittsylvaniaCo evidence: the annual county personal property tax records. A close analysis of these tax records, in the context of my TIMELINE of other PittsylvaniaCo data, the extensive USCensus abstracts that I’ve compiled over the years for all southern DENNISONS, and the ancestral pedigrees submitted by Julie-08 and Kathy-52, has enabled me to reconstruct three PittsylvaniaCo DENNISON families, headed, in order of their appearance in the county, by James, Benjamin, and Richard.
All the circumstantial indications suggest that these three were brothers, and since Kathy’s research also shows that her Benjamin came to PittsylvaniaCo from Maryland, presumably the others did too.
Of these PittsylvaniaCoVA DENNISONs, Julie-08, representing Patrilineage 2, traces back to James, while Kathy-52 of Patrilineage 6 traces back to Benjamin. Thus, despite the circumstantial evidence that indicates that these two were brothers, evidently an NPE (Non-Paternity Event) has occurred somewhere up the ancestral chain of one of them—but not downstream because both Julie and Kathy have patrilineage cousins who can trace back to sons or grandsons of one of the three PittsylvaniaCo patriarchs. It’s probable, in fact, that either James or Benjamin had a non-DENNISON father, and that these two were therefore half-brothers. If we could yDNA test a descendant of the third PittsylvaniaCo patriarch, Richard, that would presumably tell us whether James or Benjamin was the odd man odd, and therefore the patriach of the NPE line.
More research needs to be done to try to connect the as yet unconnected members of both the Patrilineage 2 and 6 projects to their respective PittsylvaniaCo ancestors, and it would be also be desirable to extend the haplotypes of these unconnected members to 111 markers. This might turn up Closer Cousin Cluster (CCC) relationships that could help guide and focus further research.
Extending should also produce more yDNA matches to LONGACREs and other surnames of the patrilineage, and give us a better reading, either on when the NPE occurred, or on the time and place that the DENNISON surname was first adopted as an alternative to the original Scandinavian name, or both. I note that FTDNA has dropped the price of extending one’s haplotype quite dramatically, and even better prices can be obtained by waiting for the several sales the company runs each year.
In the meantime, I have published all my work on this patrilineage as a set of linked reports under the heading “Other Genealogical Resources & Evidence” in the top left navigation panel of this page. You will find there: my provisional reconstruction of the family TREE of the three PittsylvaniaCo DENNISON families (with some interleaved references to the evidence that guided me); a detailed analytical report on the Three DENNISON+ Patriarchs that elucidates my thinking and backs up my conclusions in the TREE; and the various evidential compilations on which my analysis mostly rests (a PittsylvaniaCo TIMELINE; my tax list abstracts for the DENNISONs of Pittsylvania and many other VA counties; and my USCensus abstracts (possibly) pertinent to the PittsylvaniaCo DENNISONs and descendants.
At the least, my major conclusions from the tax records that James is the ancestor of Julie-08 through his son William, and that he had at least three other sons, John, James, and Stephen, should help the other unconnected members of Patrilineage 2 to particularly target possible connecting ancestors of these names.
invisible writing
4Feb2024
Welcome to the newest member of the project, James Royce Dennison, Jr. (Jim_R, Project# D-47). Jim, like existing members Julie-08, Bill_J-54, and Bob_L-26 is descended from 1-James Dennison of PittsylvaniaCoVA, but through a different son, 2-Stephen, rather than 2-William. Jim has done FTDNA’s full BigY-700 test, which should be very useful for comparison purposes if we can get one of the known Scandinavian-derived members to also order the BigY-700. Jim’s addition has also prompted me to re-evaluate and re-calibrate the association between these two very distant arms of what continues to look like a single patrilineage, although the MRCA now appears to extend centuries before surnames were assumed and recorded.
More immediately useful is that Jim’s haplotype diverges from those of the three 2-William descendants, such that we can now say for sure that two mutations, DYS390=23 and CDYb=39 (but especially the former) can be expected to brand all of the descendants of either 2-William, or of his son 3-Trammel, solidifying their status as a Closer Cousin Cluster, and Jim has also turned up a high-quality (slow to mutate) mutation of his own, DYS552=23, in the 68-111 marker band that likewise identifies his branch of descendants—except that without further testing of fellow descendants of 2-Stephen, there's no telling just how high up the ancestral tree it goes.
And on the purely genealogical side, the fact that Jim’s line can be credibly traced back to the Stephen Denison who headed a household in the 1850 USCensus for DecaturCoTN, who corresponds perfectly with the Stephen Dennison who entered into a marriage bond on 15Oct1813 in PittsylvaniaCoVA, while Jim’s haplotype corresponds so closely to those of three known descendants of the original PittsylvaniaCo settler 1-James, constitutes a pretty conclusive link between these middlewestern TN DENNISONs, and their PittsylvaniaCo place of origin. And the USCensuses of 1860 on show that several of 2-Stephen’s son subsequently migrated RedRiver and adjacent Bowie County in the NE corner of Texas.
DENNISON Patrilineage 2 yDNA Haplotypes Compared
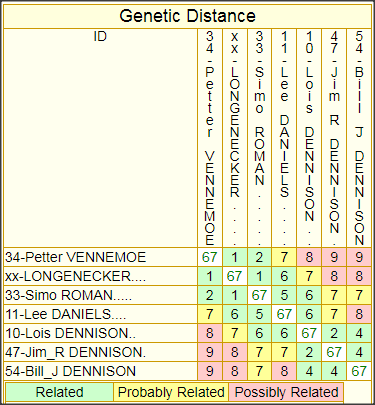
The following matrices, one for 37-marker comparisons, and one for 67-marker comparisons (which includes the ROMAN haplotype) provide some idea of the closeness of relationship between each pair of test subjects of this patrilineage. The cell at the intersection of each column/row pair shows the GD (Genetic Distance) between the pair—basically, the total number of mutations that has occurred in both of their lines of descent since their MRCA (Most Recent Common Ancestor) walked the earth.
It is also possible to create corresponding matrices that show TMRCA (Time back to the MRCA, expressed either in generations or in years), but I have decided to forgo such charts because they are just too misleading as indicators of when a particular MRCA lived. Mutations are so sporadic and infrequent (even when a large number of markers is tested) that such estimates, expressed in years, can easily be off by 100-200 years either way. If one has the urge to play around with TMRCA estimates between one’s own haplotype and those of one’s closest matches, the best way to do that is to run the built-in FTDNA Tip calculator from one’s personal page—and be sure to input the number of generations for which one knows, genealogically, that you and your match cannot have had a common ancestor.
While the GD, at least, is exact, there is no obvious way to tell how the mutations divide between the two subjects, because the haplotype of their common ancestor, from whom they have mutated, is unknown. However, I have developed a procedure using the GD chart for inferring the haplotype of the common ancestor, which I call the Root Prototype Haplotype (RPH), and I use this as a basis for marking mutations in the yDNA Haplotypes chart, elsewhere on this page.
FTDNA provides advisories for classifying GDs between haplotype pairs for their degree of closeness, or to put it otherwise, for the probability that the two have a common male ancestor within genealogical time, and thus belong to the same (genealogical) patrilineage. Unfortunately, neither FTDNA’s37-marker GD advisory, nor its 67-marker GD advisory can be relied on for an adequate elucidation of this subject. Although the latter does note the relevance of genealogical evidence, which is an improvement over the confused and misleading text of the 37-marker advisory, it too fails to note the crucial importance of two haplotypes sharing a common surname, and the too stringent Relatedness categories of the 37-marker advisory have been relaxed but only at the expense of inconsistent scaling by the respective average mutation rate of the panels.
Another important principle not made clear by the FTDNA text (although there are signs of trying) is that the relationship of an outlier to the the group is to be judged by the number and closeness of his nearest approach to it, i.e. by his lowest GD numbers, not his highest or average GD.
Finally, although the infrequent and sporadic nature of the mutation process requires that one mentally add or subtract one or two from the GD for any particular haplotype pair to get some idea of its inaccuracy as a measure, the overall structure of the matrix for a largish patrilineage tends to paint a much more accurate picture of the depth of the patrilineage as a whole. And this is particularly true for a largish matrix of 111-marker haplotypes. In fact, it is largely for this reason that I believe such matrices to be of genealogical value: not so much because they accurately indicate relationships between any particular pair of haplotypes, as because they provide a reasonably accurate idea of the relationship of all to each other, and to the MRCA that they all have in common. Even for outliers, as long as there is one closer GD relationship across the group, we may reasonably classify them as of the same patrilineage, in accordance with the principle I’ve just stated.
-
37-Marker Haplotype Genetic Distance Comparison Matrix
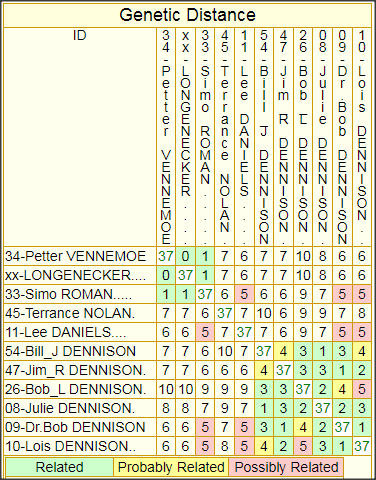
The number in each cell is the number of divergent mutations between each pair of haplotypes.
The lowest numbers represent the closest relationships.
Whether an outlier haplotype belongs to the same patrilineage should be judged by its lowest GD number at the greatest number of markers.
Color-coding shows whether a haplotype pair Definitely, Probably, or just Possibly belongs to the patrilineage.
However, the color-coded categories and the GD numbers don’t take account either of the common surname that most of these haplotypes share,
or of the possible convergence of their genealogical evidence at a particular time and place, and where either of these conditions obtain,
1 or 2 can reasonably be subtracted from the indicated GD in assessing whether an outlying haplotype belongs to the patrilineage.
-
67-Marker Haplotype Comparison Matrix 111-Marker Haplotype Comparison Matrix
-
The number in each cell is the number of divergent mutations between each pair of haplotypes.
The lowest numbers represent the closest relationships.
Whether an outlier haplotype belongs to the same patrilineage should be judged by its lowest GD number at the greatest number of markers.
Color-coding shows whether a haplotype pair Definitely, Probably, or just Possibly belongs to the patrilineage.
However, the color-coded categories and the GD numbers don’t take account either of the common surname that most of these haplotypes share,
or of the possible convergence of their genealogical evidence at a particular time and place, and where either of these conditions obtain,
1 or 2 can reasonably be subtracted from the indicated GD in assessing whether an outlying haplotype belongs to the patrilineage.
DENNISON Patrilineage 2 ySTR DNA Haplotypes
The chart below shows the haplotypes for each tested project member of this patrilineage. I’ve decapitated most of the marker names (truncating “DYS393” to just “393”) to improve readability. The markers with colored headings mutate slower or faster than the norm. Thus, [DYS]439 is a fast mutator, [DYS]458 is faster, and CDYa&b are blazing, while [DYS]393 is slow. The remaining markers fall into the midrange of mutability.
The mutability of the markers that have mutated is important because it is the shared inherited mutations that identify the Closer Cousin Clusters which it is the main purpose of testing to ascertain. When these shared mutations are to the more mutable markers there’s a fair chance that some may have mutated independently in different lines, and thus weren’t inherited from a common ancestor.
| Haplotype Identifiers | FTDNA 37-Marker Panel | FTDNA Markers 38-67 | FTDNA Markers 68-111 | Haplotype Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Proj # |
Principal Genealogist |
Earliest Known Ancestor Name (Birth - Death) c=circa s=say |
3 9 3 | 3 9 0 |
1 9 / 3 9 4 | 3 9 1 |
3 8 5 a | 3 8 5 b |
4 2 6 |
3 8 8 |
4 3 9 |
3 8 9 I |
3 9 2 |
3 8 9 I I |
4 5 8 |
4 5 9 a |
4 5 9 b |
4 5 5 |
4 5 4 |
4 4 7 |
4 3 7 |
4 4 8 |
4 4 9 |
4 6 4 a |
4 6 4 b |
4 6 4 c |
4 6 4 d |
4 6 0 |
Y G - H 4 |
Y C A I I a |
Y C A I I b |
4 5 6 |
6 0 7 |
5 7 6 |
5 7 0 |
C D Y a |
C D Y b |
4 4 2 |
4 3 8 |
5 3 1 |
5 7 8 |
3 9 5 S 1 a |
3 9 5 S 1 b |
5 9 0 |
5 3 7 |
6 4 1 |
4 7 2 |
4 0 6 S 1 |
5 1 1 |
4 2 5 |
4 1 3 a |
4 1 3 b |
5 5 7 |
5 9 4 |
4 3 6 |
4 9 0 |
5 3 4 |
4 5 0 |
4 4 4 |
4 8 1 |
5 2 0 |
4 4 6 |
6 1 7 |
5 6 8 |
4 8 7 |
5 7 2 |
6 4 0 |
4 9 2 |
5 6 5 |
7 1 0 |
4 8 5 |
6 3 2 |
4 9 5 |
5 4 0 |
7 1 4 |
7 1 6 |
7 1 7 |
5 0 5 |
5 5 6 |
5 4 9 |
5 8 9 |
5 2 2 |
4 9 4 |
5 3 3 |
6 3 6 |
5 7 5 |
6 3 8 |
4 6 2 |
4 5 2 |
4 4 5 |
Y G - A 1 0 |
4 6 3 |
4 4 1 |
Y G - 1 B 0 7 |
5 2 5 |
7 1 2 |
5 9 3 |
6 5 0 |
5 3 2 |
7 1 5 |
5 0 4 |
5 1 3 |
5 6 1 |
5 5 2 |
7 2 6 |
6 3 5 |
5 8 7 |
6 4 3 |
4 9 7 |
5 1 0 |
4 3 4 |
4 6 1 |
4 3 5 |
Proj # |
Principal Genealogist |
| D-35 | Petter Vennemoe | Svend Bøen (c1617-c1702 AkershusNOR) | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 13 | 29 | 17 | 9 | 10 | 11 | 11 | 25 | 15 | 19 | 31 | 15 | 15 | 15 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 18 | 18 | 36 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 16 | 8 | 12 | 22 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | 32 | 15 | 9 | 16 | 12 | 24 | 26 | 19 | 12 | 11 | 13 | 12 | 11 | 9 | 12 | 12 | 10 | 11 | 11 | 31 | 12 | 13 | 24 | 13 | 10 | 10 | 18 | 15 | 18 | 13 | 24 | 16 | 13 | 15 | 24 | 12 | 23 | 18 | 10 | 14 | 17 | 9 | 12 | 11 | D-35 | Petter Vennemoe |
| D-33 | Simo Roman | Anders Larsson (say 1695 SWE - [SWE]) | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 13 | 29 | 17 | 9 | 10 | 11 | 11 | 25 | 15 | 19 | 31 | 15 | 15 | 15 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 18 | 18 | 37 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 22 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | D-33 | Simo Roman | ||||||||||||||||||||||||||||||||||||||||||||
| LONGENECKER | Anders Larsson (say 1695 SWE - [SWE]) | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 13 | 29 | 17 | 9 | 10 | 11 | 11 | 25 | 15 | 19 | 31 | 15 | 15 | 15 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 18 | 18 | 36 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 22 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | 33 | 15 | 9 | 16 | 12 | 24 | 26 | 19 | 12 | 11 | 13 | 12 | 11 | 9 | 12 | 12 | 10 | 11 | 11 | 31 | 12 | 13 | 24 | 13 | 10 | 10 | 19 | 15 | 20 | 13 | 24 | 16 | 13 | 15 | 24 | 12 | 23 | 18 | 10 | 14 | 17 | 9 | 12 | 11 | LONGENECKER | ||
| D-11 | Lee Daniels | ThomasDaniels (c1762-1828 Bucks.ENG) | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 13 | 29 | 17 | 9 | 10 | 11 | 11 | 24 | 15 | 19 | 30 | 14 | 15 | 15 | 17 | 10 | 12 | 19 | 23 | 17 | 15 | 20 | 18 | 37 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 22 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | D-11 | Lee Daniels | ||||||||||||||||||||||||||||||||||||||||||||
| RPH: Scandinavia | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 13 | 29 | 17 | 9 | 10 | 11 | 11 | 25 | 15 | 19 | 31 | 15 | 15 | 15 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 19 | 18 | 37 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 22 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | 33 | 15 | 9 | 16 | 12 | 24 | 26 | 19 | 12 | 11 | 13 | 12 | 11 | 9 | 12 | 12 | 10 | 11 | 11 | 31 | 12 | 13 | 24 | 13 | 10 | 10 | 19 | 15 | 18 | 13 | 24 | 16 | 13 | 15 | 24 | 12 | 23 | 18 | 10 | 14 | 17 | 9 | 12 | 11 | RPH | |||
| RPH: Virginia | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 12 | 29 | 17 | 9 | 10 | 11 | 11 | 25 | 15 | 19 | 32 | 15 | 15 | 17 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 19 | 18 | 37 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 21 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | 34 | 15 | 9 | 16 | 12 | 24 | 26 | 19 | 12 | 11 | 13 | 12 | 11 | 9 | 12 | 12 | 10 | 11 | 11 | 31 | 12 | 13 | 24 | 13 | 10 | 10 | 19 | 15 | 18 | 13 | 24 | 17 | 12 | 16 | 24 | 12 | 23 | 18 | 10 | 14 | 17 | 9 | 12 | 11 | RPH | |||
| D-10 | Lois Franceschi | Joel (c1808 VA -) | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 12 | 29 | 17 | 9 | 10 | 11 | 11 | 24 | 15 | 19 | 32 | 15 | 15 | 17 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 19 | 18 | 37 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 21 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | D-10 | Lois Franceschi | ||||||||||||||||||||||||||||||||||||||||||||
| D-09 | Dr. Bob Dennison | Stephen (c1789 VA -) | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 12 | 29 | 17 | 9 | 10 | 11 | 11 | 24 | 15 | 19 | 33 | 15 | 15 | 17 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 19 | 18 | 37 | 38 | 12 | 12 | D-09 | Dr. Bob Dennison | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D-08 | Julie Dennison | James (say 1752 MD? - PittsylvaniaCoVA) | 13 | 23 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 12 | 29 | 17 | 9 | 10 | 11 | 11 | 24 | 15 | 19 | 33 | 15 | 15 | 17 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 19 | 18 | 37 | 39 | 12 | 12 | D-08 | Julie Dennison | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D-54 | William J. Dennison | James (say 1752 MD? - PittsylvaniaCoVA) | 13 | 23 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 12 | 29 | 17 | 9 | 10 | 11 | 11 | 24 | 15 | 19 | 33 | 15 | 15 | 17 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 18 | 18 | 37 | 39 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 21 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | 34 | 15 | 9 | 16 | 12 | 24 | 26 | 19 | 12 | 11 | 13 | 12 | 11 | 9 | 12 | 12 | 10 | 11 | 11 | 31 | 12 | 13 | 24 | 13 | 10 | 10 | 19 | 15 | 18 | 13 | 24 | 17 | 12 | 16 | 24 | 12 | 23 | 18 | 10 | 14 | 17 | 9 | 12 | 11 | D-54 | William J. Dennison |
| D-26 | Robert L. Dennison | James (say 1752 MD? - PittsylvaniaCoVA) | 13 | 23 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 12 | 29 | 17 | 9 | 10 | 11 | 11 | 24 | 15 | 19 | 33 | 15 | 15 | 17 | 17 | 10 | 11 | 19 | 23 | 18 | 15 | 19 | 19 | 37 | 39 | 12 | 12 | D-26 | Robert L. Dennison | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D-47 | James R. Dennison | Stephen (c1789 VA - say 1855 TN) | 13 | 24 | 14 | 10 | 11 | 14 | 12 | 12 | 12 | 13 | 12 | 29 | 17 | 9 | 10 | 11 | 11 | 24 | 15 | 19 | 33 | 15 | 15 | 17 | 17 | 10 | 11 | 19 | 23 | 17 | 15 | 19 | 19 | 37 | 38 | 12 | 12 | 11 | 9 | 15 | 16 | 8 | 11 | 10 | 8 | 10 | 10 | 12 | 23 | 23 | 16 | 10 | 12 | 12 | 15 | 8 | 12 | 21 | 21 | 14 | 12 | 11 | 13 | 11 | 11 | 12 | 12 | 34 | 15 | 9 | 16 | 12 | 24 | 26 | 19 | 12 | 11 | 13 | 12 | 11 | 9 | 12 | 12 | 10 | 11 | 11 | 31 | 12 | 12 | 24 | 13 | 10 | 10 | 19 | 15 | 18 | 13 | 24 | 17 | 12 | 16 | 23 | 12 | 23 | 18 | 10 | 14 | 17 | 9 | 12 | 11 | D-47 | James R. Dennison |
You may click on highlighted Project#s (like D-08) to see the posted pedigree for a particular test subject. Click on highlighted Researcher names, like Julie Dennison to go to the project directory that shows the full names of the members, and provides clickable e-mail links for the names highlighted there.
The chart scrolls to the right to show tested markers beyond the first 37. A synthetic Root Prototype Haplotype (RPH) has been constructed to represent the most likely (unmutated) haplotype of the Most Recent Common Patriarchal Ancestor (MRCPA) of all the members of this project. In most cases the marker values of this RPH are those which are the most common across all of the haplotypes. Marker values that deviate from those of the RPH are deemed to be mutations, and are highlighted in lime green— or in tomato for possible multistep mutations, though some of these may have mutated more than once. Markers with null values, due to deletions, are rendered in dark seagreen. Where the multicopy markers DYS464 and YCA (each taken as a whole) diverge in value from those of the RPH, the whole adjacent set of values will be colored yellow green, and will be counted as a single mutation. In the same way, reclOH mutations, which may affect several blocks of separated markers, will be colored orange and treated all as a single mutation when calculating Genetic Distance.
You may click on a highlighted Project Member # (like D-26 for member Bob_L) to see the posted pedigree for that particular test subject. Clicking on the name of the Principal Genealogist, like Robert L. Denison brings up his entry in the Directory of Researchers, and clicking on his full name in the Directory brings up an email blank addressed to him. The Earliest Known Patrilineal Ancestor (and/or his data) is as projected by the editor (JBR), and may not reflect the opinion of the Principal Genealogist of that line.
*** THIS CONTENT WAS STOLEN FROM JOHNBROBB.COM IN VIOLATION OF COPYRIGHT LAW ***
Last updated 4Feb2024
© John Barrett Robb


*** THIS PAGE WAS STOLEN FROM JOHNBROBB.COM IN VIOLATION OF COPYRIGHT LAW ***